神经系统特异标记的转基因斑马鱼品系推荐
作者:谢训卫 发布时间:2019/10/10 5:00:00斑马鱼是研究神经系统发育、功能和组织修复的理想模型,尤其是特异性表达荧光蛋白的转基因品系,帮助我们标记目标组织和细胞,可以清晰直观地观察到活体神经细胞的动态生物学过程。
国家斑马鱼资源中心现有各类研究用品系1400多种,其中转基因品系近200种。本文为大家推荐其中常用的几种神经系统标记品系。
.jpg)
双色荧光标记的斑马鱼神经系统
斑马鱼中枢神经系统自前段向后依次为:嗅球(olfactory bulb)、端脑(telencephalon)、间脑(diencephalon)、下丘脑(hypothalamus)、视顶盖(optic tectum)、小脑(cerebellum)、后脑(rhombencephalon)和延髓(medulla)等基本结构,如下图所示。幼年期的斑马鱼大脑组织透明,且结构简单,可以作为优良的神经系统动物模型(Shang et al. 2015)。
.jpg)
斑马鱼神经系统 (Grandel et al. 2006)
能全面标记中枢神经系统的品系首推CZ417(ihb304Tg, Tg(gad1b:mCherry)),该品系由gad1b启动子驱动红色荧光蛋白mCherry的表达。gad1b是编码生成谷氨酸脱羧酶(Glutamic Acid Decarboxylase,GAD)亚型GAD67的基因。GAD负责催化谷氨酸(glutamate)的脱羧反应,产生γ-氨基丁酸(Gamma Amino Butyric Acid,GABA)和CO2。GABA是起抑制性作用的神经递质,因此gad1b启动子通常是作为抑制性神经元的特异性启动子。gad1b基因的表达谱很广,从嗅球(olfactory bulb)、视顶盖(optic tectum)、眼睛(eye)、延髓(medulla oblong)和脊髓(spinal cord)都可以观察到红色荧光蛋白的表达(Song et al., 2017)。
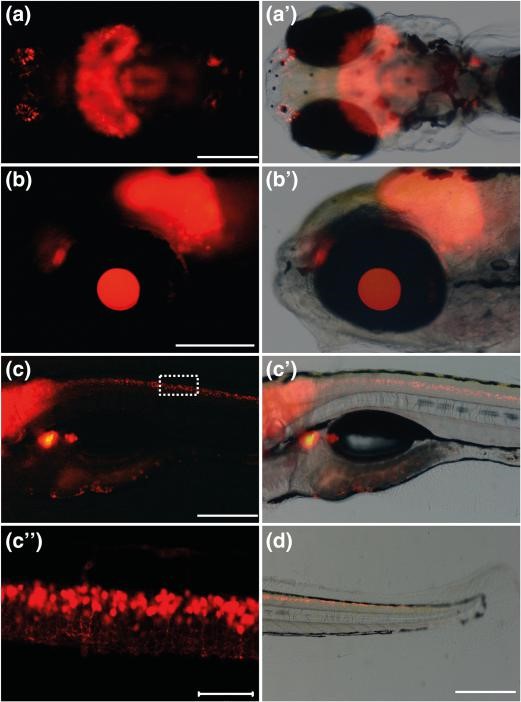
CZ417, ihb304Tg, Tg(gad1b:mCherry) (Song et al., 2017)
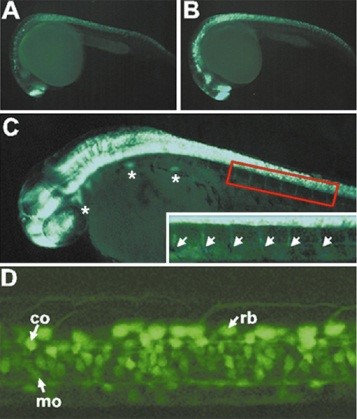
CZ413(tsu33Tg, Tg1(elavl3:YFP))和CZ160(knu3Tg, Tg(elavl3:EGFP))分别使用神经发育标记基因elavl3的启动子驱动YFP和EGFP的表达,在全身的神经元中特异表达标记基因。elavl3基因别名huc,最早在果蝇中发现,该基因缺失会导致视觉神经系统缺陷伴随神经系统发育不良,严重致死。刚过原肠期,Elavl3基因开始在分布在中线两侧的神经元中表达,神经系统二次发育开始不久,Elavl3基因表达急剧上升。elavl3在斑马鱼早期神经发育过程中发挥了重要的作用,是神经系统发育的重要标记基因(Lyons et al., 2003; Zhao et al., 2006; Wu et al., 2018)。
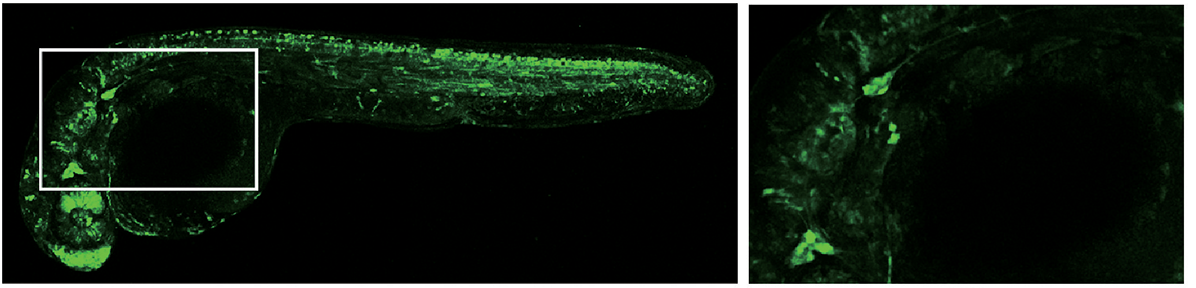
CZ413, tsu33Tg, Tg1(elavl3:YFP)( Wu et al., 2018)

CZ413, tsu33Tg, Tg1(elavl3:YFP)
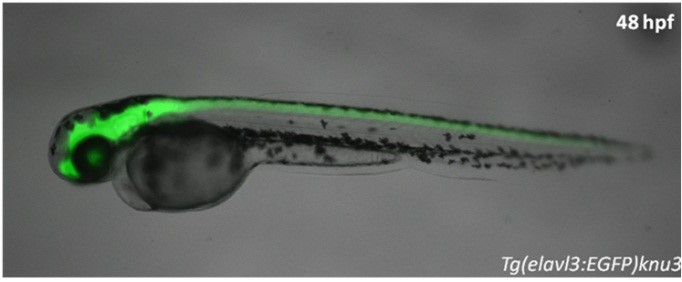
CZ160, knu3Tg, Tg(elavl3:EGFP)
CZ154(vu19Tg, Tg(olig2:DsRed2))转基因利用红色荧光蛋白DsRed2标记运动神经元和头部少突胶质细胞群。在神经系统发育过程中,放射状胶质细胞既是神经元的前身,也充当着新生神经元迁移的支架。Notch信号通路在放射状胶质细胞的维持中发挥了重要的作用,同时抑制其形成运动神经元,促进其向少突胶质细胞分化(Kim et al., 2008)。放射状胶质细胞在发育中的中枢神经系统中居要角,扮演引导神经元迁移的角色。缺少胶质细胞的胚胎中,运动神经元会偏离预定迁移轨迹,向脊髓外迁移,形成异常的轴突投射(Kucenas et al., 2008)。
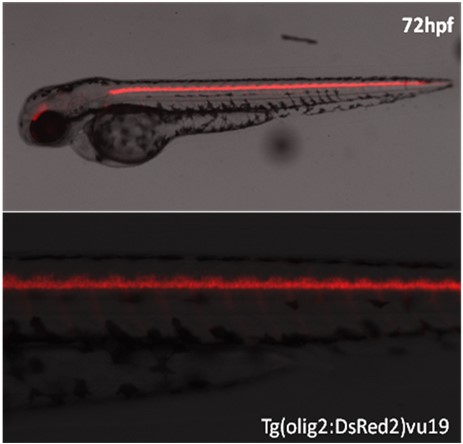
CZ154, vu19Tg, Tg(olig2:DsRed2)
CZ224(nju1Tg, Tg(cyp26a1:EYFP))转基因品系由cyp26a1基因一段2.5kb的启动子驱动EYFP标记基因的表达,表达部位主要分布在视网膜、嗅球、背侧脊髓前部、后肠、脊索后部和咽弓等组织中(Hu et al., 2008)。cyp26a1基因是细胞色素P450家族成员, 相对应的产物为Cyp26氧化酶,可以和视黄醛脱氢酶RALDHs(aldh1a2基因产物)共同协调维持视黄酸(RA)的均衡状态。视黄酸参与了细胞增殖和凋亡、细胞分化和组织形态等多个重要的生理功能中,对机体的视觉、皮肤、免疫、生殖和神经系统等都有一定的影响。
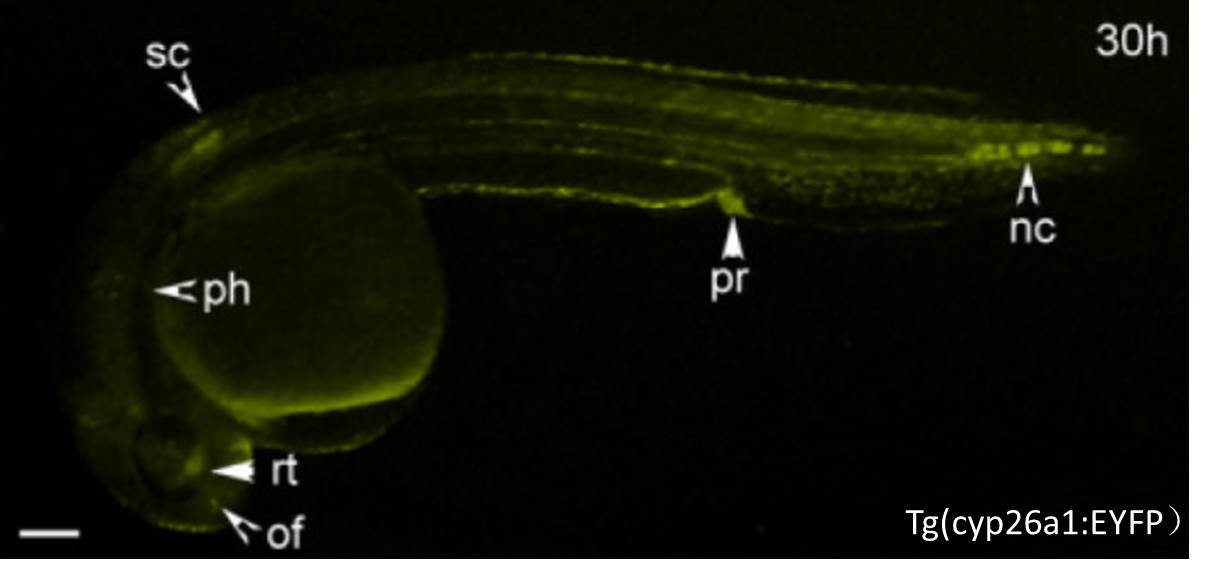
CZ224, nju1Tg, Tg(cyp26a1:EYFP)
对特异单一神经相关组织感兴趣的朋友可以关注CZ25(Tg(OTM:dGFP) ihb27Tg,标记中脑组织),CZ31(Tg(-1.0pomca:GFP) zf44Tg,标记脑垂体),CZ36(Tg(-2.5tshb:EGFP) ihb5Tg,标记松果体),CZ151(Tg(ompb:tau-eGFP) jt0012Tg,标记嗅球),CZ153(Tg(mbp:EGFP) ck1Tg,标记成髓鞘胶质细胞),CZ353(Tg(atoh1a:dTomato)nns8Tg,标记小脑组织)和CZ415(Tg(crybb1:CFP) tsu24Tg,标记晶状体)等品系。
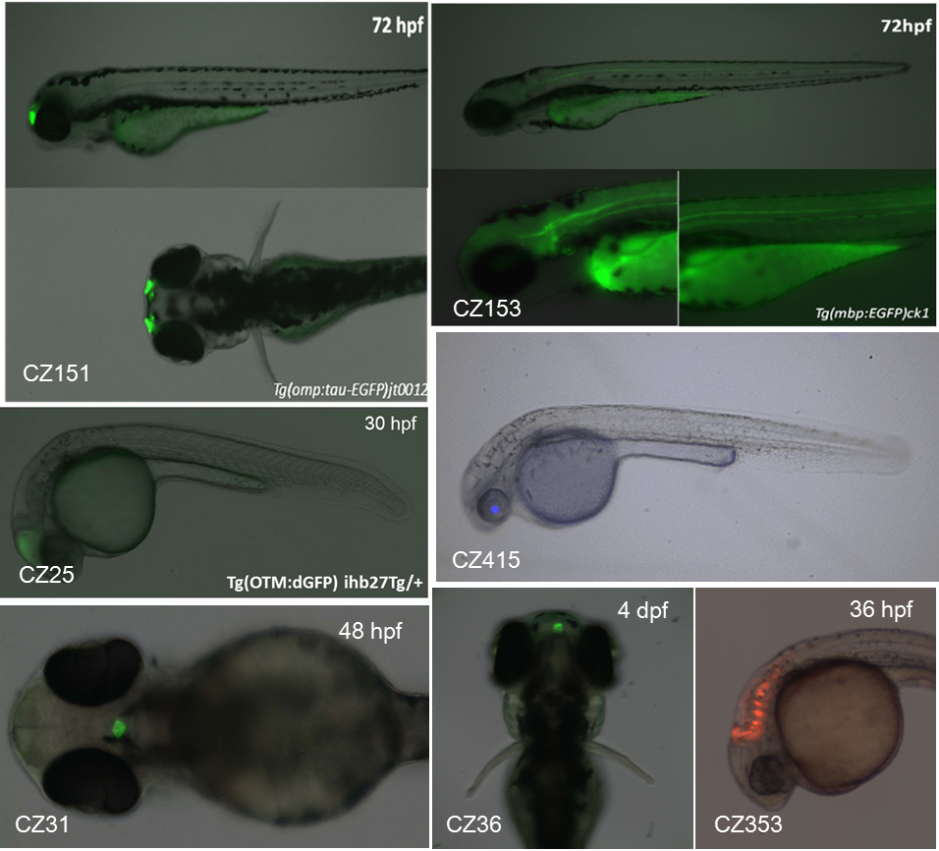
标记单一特异神经组织的品系
还有很多有意思的品系在这里就不一一赘述了,有兴趣的朋友请点击品系列表的CZ号就可以进入品系信息页面了解啦。未来中心的全体员工将继续努力以为斑马鱼研究团队提供更好的科研支撑。
|
CZ ID |
Genotype |
construct |
Note |
|
ihb27Tg/+ |
Tg(OTM:GFP) |
Optimal Tcf motif, TOP driving GFP expression, midbrain GFP |
|
|
zf44Tg/+ |
Tg(-1.0pomca:GFP) |
pituitary |
|
|
ihb5Tg/+ |
Tg(-2.5tshb:EGFP) |
pituitary/pineal/thyroid |
|
|
Tg(atoh7:Gal4); Tg(UAS:mcherry) |
Tg(atoh7:Gal4); Tg(UAS:mcherry) |
retinal ganglion cells |
|
|
cf2Tg/+ |
Tg(elavl3:YC2) |
neurons, retina |
|
|
ml3Tg/+ |
Tg(mnx1:mGFP) |
Primary motor axons, motoneurons |
|
|
rw0Tg/+ |
Tg(isl1:GFP) |
cranial motor neuron |
|
|
zf147Tg/+ |
Tg(apoeb:LY-EGFP) |
Neuronal |
|
|
kca66Tg/+ |
Tg(h2afva:h2afva-GFP) |
neuromast primordium migration, posterior lateral line nerve development |
|
|
ck2Tg/+ |
Tg(eef1a1l1:Kaede) |
Brain, hatching gland |
|
|
jt0012Tg/+ |
Tg(ompb:tau-EGFP) |
Olfactory bulb, Olfactory placode |
|
|
ck1Tg/+ |
Tg(mbp:EGFP) |
myelinating glia, central nervous system |
|
|
vu19Tg/+ |
Tg(olig2:DsRed2) |
Oligodendrocyte, motor neuron |
|
|
Tg(mbp:mGFP) |
Tg(mbp:mGFP) |
myelinating glia |
|
|
knu3Tg/+ |
Tg(elavl3:EGFP) |
neurons, nervous system |
|
|
Tg(ift46:GAL4-VP16;UAS:nfsB-mCherry) |
Tg(ift46:GAL4-VP16;UAS:nfsB-mCherry) |
eye, pronephric duct, spinal cord |
|
|
nju1Tg/+ |
Tg(cyp26a1:EYFP) |
retina,olfactory vesicle, anterior dorsal spinal cord,proctodeum,caudal notochord,pharyngeal arches |
|
|
nns8Tg/+ |
Tg(atoh1a:dTomato) |
cerebellum |
|
|
nl1Tg/+ |
TgBAC(neurod1:EGFP) |
Cranial ganglia, lateral line nerve cells |
|
|
um14Tg/+ |
Tg(EPV.Tp1-Mmu.Hbb:EGFP) |
Notch-responsive tissues such as the developing CNS, vasculature, liver, intestine and pancreas |
|
|
Tg(huc:YFP)/+ |
Tg1(elavl3:YFP) |
neurons |
|
|
Tg(crybb1:CFP)/+ |
Tg(crybb1:CFP) |
eye lens |
|
|
ihb304Tg/+ |
Tg(gad1b:mCherry) |
olfactory pit, optic tectum, medulla oblong, eye, spinal cord |
神经系统特异表达的转基因斑马鱼品系(直接点接CZ号即可进入相应品系信息页面)
参考文献
1.Shang C F, Mu Y, Du J L. Zebrafish swimming into neuroscience research: a visible mind in a transparent brain. SCIENTIA SINICA Vitae, 2015, 45: 223–236, doi: 10.1360/N052014-00203
2.Grandel H, Kaslin J, Ganz J, et al. Neural stem cells and neurogenesis in the adult zebrafish brain: origin, proliferation dynamics, migration and cell fate. Developmental biology, 2006, 295(1): 263-277.
3.Song, Y., Tao, B., Chen, J., Jia, S., Zhu, Z., Trudeau, V.L., Hu, W. (2017) GABAergic Neurons and Their Modulatory Effects on GnRH3 in Zebrafish. Endocrinology. 158(4):874-886
4.Obholzer N, Wolfson S, Trapani J G, et al. Vesicular glutamate transporter 3 is required for synaptic transmission in zebrafish hair cells[J]. Journal of Neuroscience, 2008, 28(9): 2110-2118.
5.Wu X, Shen W, Zhang B, et al. The genetic program of oocytes can be modified in vivo in the zebrafish ovary[J]. Journal of Molecular Cell Biology, 2018, 10(6): 479-493.
6.Zhao C, He X, Tian C, et al. Two GC-rich boxes in huC promoter play distinct roles in controlling its neuronal specific expression in zebrafish embryos[J]. Biochemical and biophysical research communications, 2006, 342(1): 214-220.
7.Kucenas S, Takada N, Park H C, et al. CNS-derived glia ensheath peripheral nerves and mediate motor root development[J]. Nature neuroscience, 2008, 11(2): 143.
8.Kim H, Shin J, Kim S, et al. Notch‐regulated oligodendrocyte specification from radial glia in the spinal cord of zebrafish embryos[J]. Developmental dynamics: an official publication of the American Association of Anatomists, 2008, 237(8): 2081-2089.
9.Lyons D A, Guy A T, Clarke J D W. Monitoring neural progenitor fate through multiple rounds of division in an intact vertebrate brain[J]. Development, 2003, 130(15): 3427-3436.
10.Hu, P., Tian, M., Bao, J., Xing, G., Gu, X., Gao, X., Linney, E., and Zhao, Q. (2008) Retinoid regulation of the zebrafish cyp26a1 promoter. Dev. Dyn. 237(12):3798-3808