华南理工大学张文清/刘伟团队揭示Rb1在神经元凋亡程序激活和调节中的具体作用和分子机制
作者:CZRC 发布时间:2024/4/29 11:00:00
神经板的神经外胚层产生神经上皮细胞(NECs,也称为神经干细胞),从而形成神经前体,分化为组成中枢神经系统(CNs)的各种神经元和胶质细胞。神经干细胞/前体细胞(NSPCs)位于离管腔最近的心室区,在胚胎早期神经发生过程中可以快速增殖产生大量过剩的神经细胞。随后,大约50%的神经细胞在神经系统成熟之前通过凋亡清除。这种凋亡过程对于维持正常组织大小的稳态和去除-不再需要的瞬时功能细胞,以及确保每个大脑区域都有适当数量的神经元和神经胶质细胞非常重要。然而,一旦神经元成熟,它们的凋亡能力就会受到限制,从而使成熟的神经系统在一生中保持健康和功能状态。在某些病理情况下,成熟神经元的凋亡途径可以被重新激活。
神经退行性疾病影响数百万人,导致残疾和死亡。尽管神经退行性疾病(帕金森病(PD)、阿尔茨海默病(AD)、亨廷顿病(HD)和肌萎缩性侧索硬化症(ALS)等)在不同神经元亚群的病理遗传改变和变性方面存在差异,但许多研究已经提供了这些疾病中凋亡通路激活(如Caspase-3/8/9、Bax过表达和Bcl-2减少)的证据。此外,越来越多的研究表明,在AD、PD和ALS中,存在细胞周期相关蛋白增加神经元死亡的风险,提示神经元细胞周期再进入参与了神经元的凋亡和死亡。有趣的是,细胞周期负调控因子RB1的过度磷酸化与神经退行性疾病有关。研究人员在AD患者、ALS患者和PD患者的病理组织中检测到RB1的过度磷酸化。这些研究表明,RB1可能参与神经元凋亡,但没有直接的体内证据证明RB1在这一过程中的作用,并且细胞凋亡途径的潜在机制仍有待揭示。
2024年4月18日,华南理工大学医学院张文清/刘伟团队在学术期刊Cell Death Discovery上发表了题为“Role of RB1 in neurodegenerative diseases: inhibition of post-mitotic neuronal apoptosis via Kmt5b”的文章,该研究揭示了Rb1在神经元凋亡程序激活和调节中的具体作用和分子机制,为Rb1相关神经系统疾病的发病机制提供参考。

已有研究报道发现神经退行性患者(如AD、ALS和PD)的病理组织中RB1的过度磷酸化。为了确定RB1突变与神经退行性疾病之间的关系,我们使用金域医学检验集团股份有限公司的DNA测序数据计算了PD(189份血样)、AD(33份血样),HD(69份血样)和ALS(75份血样)的RB1突变率。结果显示,RB1突变的频率在PD中为2.64%,在AD中为3.03%,在HD中为2.85%,在ALS中为2.66%(图1A)。这些RB1突变存在于9名神经退行性疾病患者中,包括6个位点的突变(4个错义突变位点:G38D、R621S、R798W和L819V;2个剪接突变位点:X405和X406)(图1B和C)。神经退行性疾病患者中四个错义等位基因的频率略高于ALAF项目和来自NCBI的正常东亚人群,但只有R621S和L819V突变显示出统计学意义(图1C)。总之,通过分析临床测序数据,我们发现RB1突变在神经退行性疾病中占2-3%。
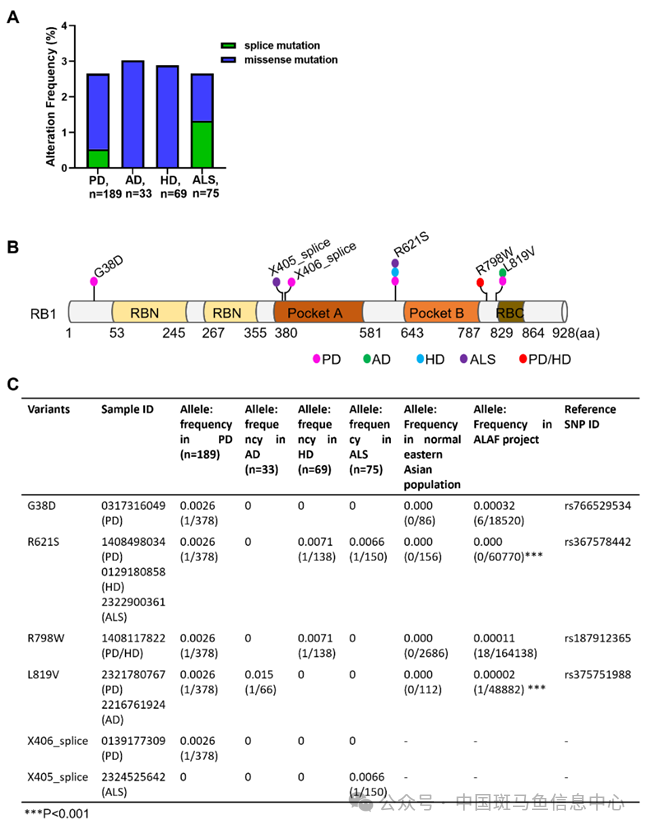
图1 神经退行性疾病患者中RB1发生改变
图1 神经退行性疾病患者中RB1发生改变
神经退行性疾病的特点是进行性认知功能障碍和行为异常。为了进一步了解Rb1在这些疾病中的作用,我们在斑马鱼中获得了一个Rb1突变体(命名为zrb1 KO)。zrb1 KO突变体在外显子2中有2个碱基缺失,这产生了一个过早的终止密码子,并显著降低了rb1 RNA的表达水平。用行为轨迹跟踪系统研究了zrb1-/-幼鱼的游泳行为,用T迷宫研究了zrb1+/-成鱼的认知能力。我们的结果显示,zrb1-/-幼鱼的游泳距离和速度显著降低(图2A),表明Rb1缺失导致幼鱼运动功能障碍。由于zrb1-/-(在约15 dpf时死亡)的胚胎致死性,我们使用zrb1+/-成鱼(3个月)来测试它们的游泳行为和认知能力。结果显示,zrb1+/-成鱼的游泳距离和速度正常,但他们的空间学习和记忆能力下降(图2B)。在食物刺激训练之前,wt和zrb1+/-成鱼在左臂和右臂停留时间没有差异。然而,在右臂(富集室:EC)中进行7天的食物刺激训练后,wt在右臂中的停留时间增加,而zrb1+/-成鱼在左臂和右臂停留时间没有差异(图2B)。这些发现表明,Rb1缺乏会损害斑马鱼幼鱼的运动能力和杂合子成鱼的认知能力。
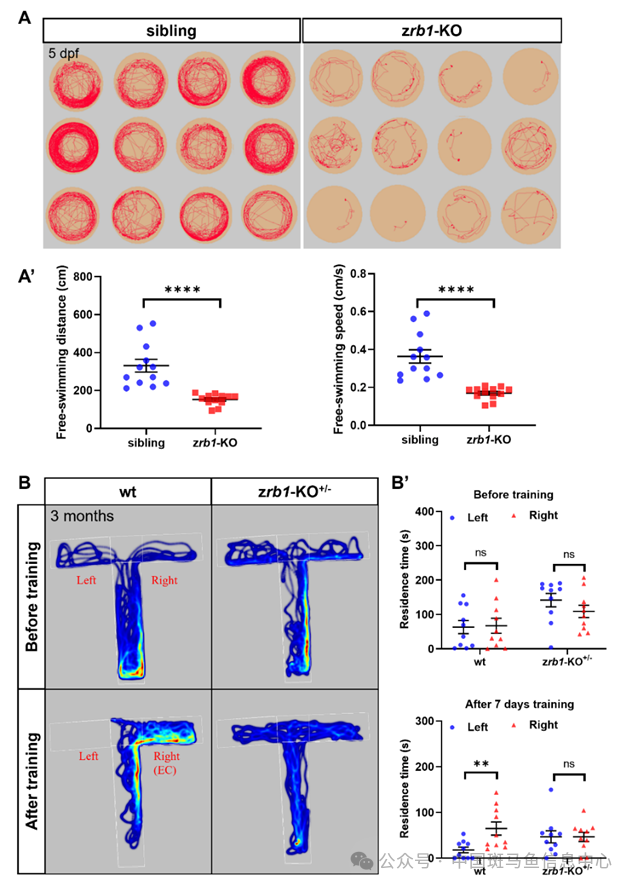
图2 Rb1缺失会导致幼年斑马鱼的游泳行为和成年斑马鱼的记忆认知功能障碍
图2 Rb1缺失会导致幼年斑马鱼的游泳行为和成年斑马鱼的记忆认知功能障碍
由于细胞凋亡是广泛的神经退行性疾病的一个显著特征,我们研究了Rb1在神经元凋亡中的作用。通过共聚焦的亮场成像,我们观察到zrb1 KO突变体后脑中凋亡小泡的显著增加(图3A)。小胶质细胞是大脑的先天免疫细胞,可以集中在神经元死亡的区域,以消除凋亡细胞碎片。因此,我们使用中性红(NR)染色标记小胶质细胞,并观察到小胶质细胞在zrb1 KO突变体的小脑和延脑中的扩张和浸润(图3B)。与这些发现一致,AO染色显示zrb1 KO突变体的小脑和延脑中神经元凋亡增加(图3C)。为了进一步验证rb1缺失诱导后脑神经元凋亡是否是一种自主效应,我们将野生型zrb1 mRNA注射到zrb1 KO胚胎中。结果显示,在zrb1 KO突变体中注射zrb1 mRNA可以部分挽救小脑和延脑中的神经元凋亡(图3D),表明zrb1 KO突变体中细胞凋亡的增加确实是由于rb1基因的缺失引起的。总之,这些发现表明,Rb1缺陷突变体表现出显著的后脑神经元凋亡和小胶质细胞后脑浸润增加,野生型zrb1 mRNA可以部分挽救这些表型。
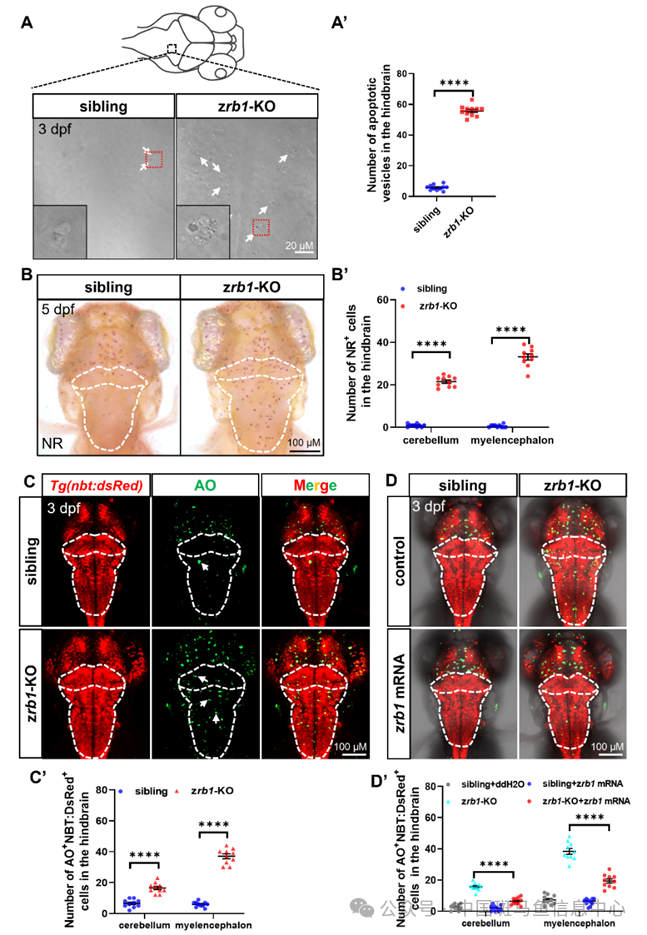
图3 rb1缺失诱导后脑神经元凋亡增加
图3 rb1缺失诱导后脑神经元凋亡增加
为了表征zrb1 KO突变体中凋亡神经元类型,我们对3 dpf的sibling和zrb1 KO突变体全脑细胞进行了10×单细胞RNA测序(scRNA-seq)。基于均匀流形近似和投影(UMAP)分析,我们对四种主要细胞类型(视神经元、NSPCs、有丝分裂后神经元和非神经元细胞)进行了聚类和注释,得到28个群。随后,将NSPCs和有丝分裂后神经元群重新聚类为六个亚群(NSPCs、前脑神经元、中脑神经元、小脑神经元、延脑神经元等),包括22个群(图4A)。在sibling和zrb1 KO突变体之间的亚群中进行了KEGG途径富集的差异表达基因(DEG)分析。有趣的是,结果显示,凋亡途径在有丝分裂后延脑神经元和小脑神经元的DEG中富集(图4B和C),而细胞周期途径在NSPCs群体中富集。这些数据表明,Rb1的缺失诱导斑马鱼后脑有丝分裂后神经元凋亡以及NSPCs的增殖。为了进一步阐明Rb1调节增殖和凋亡的细胞群的差异,我们修饰并获得了两个质粒nestin:-Cas9-T2A-mCherry,U6:gRNA(rb1)和huc:Cas9-T2Am-Cherry,U6:gRNA(rb1),其特异性敲低了NSPCs和有丝分裂后神经元中的rb1。结果显示,抑制有丝分裂后神经元中的rb1表达仅诱导小脑和延脑的细胞凋亡(图4D),而抑制NSPCs中的rb1表达诱导小脑和延脑的细胞增殖,但不诱导细胞凋亡(图4E)。
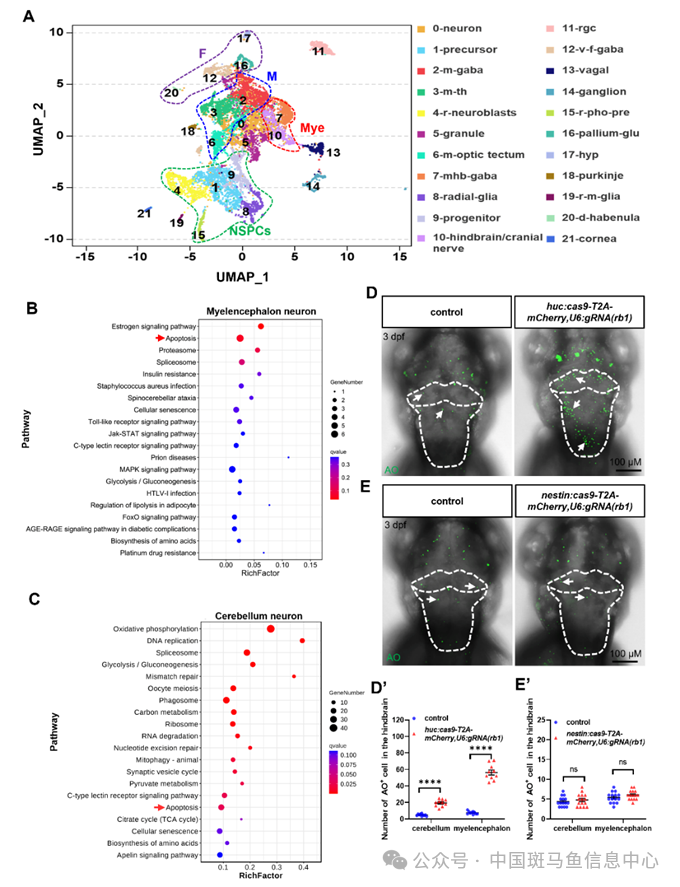
图4 Rb1调节有丝分裂后神经元的凋亡
总之,这些数据表明,zrb1 KO突变体中的凋亡细胞是后脑有丝分裂后神经元,Rb1调节有丝分裂前神经元的凋亡与其对NSPCs增殖的影响无关。
图4 Rb1调节有丝分裂后神经元的凋亡
先前的研究表明,Rb1失活后释放的E2F家族成员(主要是E2f1、E2f2和E2f3)可以激活ARF、TAp73和caspase,并通过p53依赖性或非依赖性途径诱导癌症细胞凋亡。为了阐明Rb1在小脑和延脑神经元中的凋亡途径,我们在zrb1 KO突变体中过表达bcl2,并且我们还用pan-caspase抑制剂(Z-VAD-FMK)处理zrb1 KO突变体,以抑制caspase通路。然后,通过AO染色检测凋亡细胞的数量。结果显示,在zrb1 KO突变体中bcl2a的过表达或caspase的抑制降低了小脑和延脑中凋亡细胞的数量(图5A和B),表明Rb1对神经元凋亡的调节主要取决于Bcl2和caspase通路。
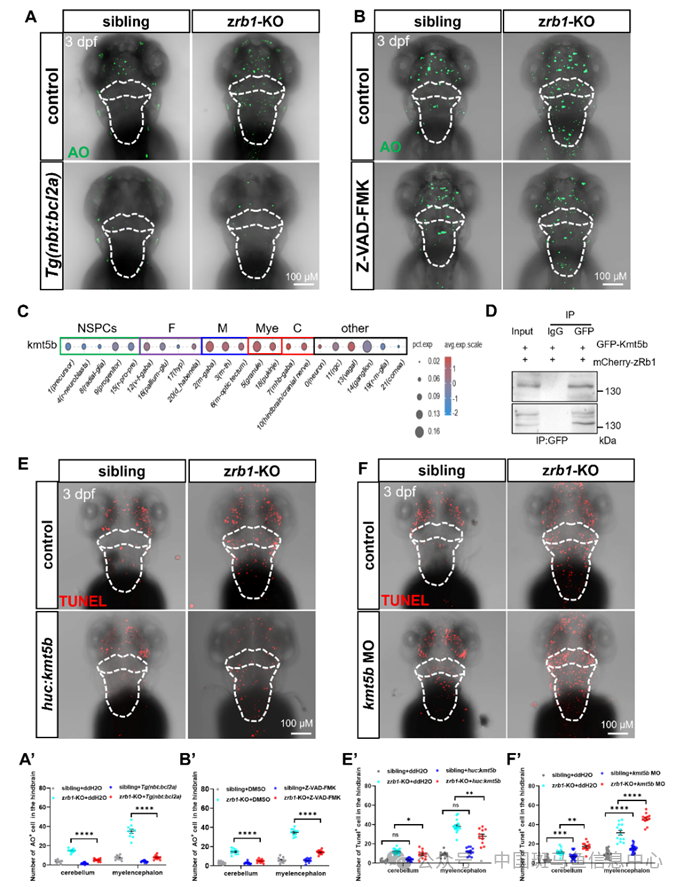
图5 Rb1通过Kmt5b-bcl2a/caspase途径调节有丝分裂后神经元凋亡
图5 Rb1通过Kmt5b-bcl2a/caspase途径调节有丝分裂后神经元凋亡
我们使用scRNA-seq数据分析了Rb1的已知结合蛋白(如E2Fs、Hdac1、Kmt5b、Dnmt1等)的表达。结果显示,只有Kmt5b(Rb1结合蛋白可以被Rb1增强)在小脑和延脑的有丝分裂后神经元中高表达(图5C)。研究表明,Kmt5b与神经发育和智力发育障碍有关。为了研究Kmt5b是否参与RB1调节的神经元凋亡途径,我们在斑马鱼中验证了RB1和Kmt5b之间的相互作用。我们在wt斑马鱼中共表达全长mCherry-zRb1和GFP–Kmt5b,并进行免疫-沉淀实验。结果表明,Rb1确实可以与斑马鱼体内的Kmt5b相互作用(图5D)。此外,我们在有丝分裂后神经元启动子(huc)下过表达kmt5b(huc:kmt5b-egfp)或在sibling和zrb1 KO突变体中通过MO敲低kmt5b观察凋亡。结果显示,在zrb1 KO突变体中过表达kmt5b部分挽救了小脑和延脑的细胞凋亡(图5E),而敲低kmt5b部分模拟了zrb1 KO突变体的表型(图5F)。同时,敲低kmt5b导致zrb1 KO突变体中bcl2a的表达降低,casp3和casp9的表达增加。这些数据表明,Rb1可以结合Kmt5b抑制bcl2a/caspase的表达,并调节凋亡途径以维持有丝分裂后神经元的存活。
斑马鱼突变体是临床实践中快速评估临床意义未明变异(VUS)的有力工具。神经元细胞死亡在许多神经退行性疾病中发挥作用。因此,我们使用zrb1 KO突变体来验证人类RB1中R621S/L819V改变(在神经退行性疾病中具有统计学意义的突变,图1C)对神经元凋亡的影响。结果表明,将人RB1 mRNA注射到zrb1 KO突变体中可以部分挽救小脑和延脑中的神经元凋亡表型(图6A),而人RB1R621S mRNA或RB1L819V mRNA不能改变zrb1 KO突变体中小脑和延脑中凋亡细胞的数量(图6A)。为了证明hRB1R621S和hRB1L819V突变是否消除RB1和Kmt5b之间的相互作用,我们在293T细胞中进行免疫沉淀测定。结果表明,hRB1可以与Kmt5b相互作用,而hRB1R621S和hRB1L819V突变不能与Kmt5b相互作用(图6B)。这表明hRB1R621S和hRB1L819V突变可能消除RB1和Kmt5b之间的相互作用,导致zrb1 KO中凋亡表型的无法挽救。上述结果表明,人RB1的R621S和L819V突变可能在神经元凋亡中发挥作用,并与神经退行性疾病有关。

图6 RB1的R621S和L819V突变可能在神经元凋亡中起作用
研究者认为这一发现揭示了Rb1在神经元凋亡程序激活和调节中的特异性作用和分子机制,并提供了一个Rb1相关神经系统疾病的发病机制参考。
华南理工大学医学院赵爽博士为本文的第一作者,张文清教授,刘伟副教授并列通讯作者。华南理工大学医学院为第一作者单位。本课题受到国家重点研发(2018YFA0801000)、广东省基础与应用基础研究基金(2023A1515010396)和广州市科技计划(202201010156)项目资助。
图6 RB1的R621S和L819V突变可能在神经元凋亡中起作用
华南理工大学医学院赵爽博士为本文的第一作者,张文清教授,刘伟副教授并列通讯作者。华南理工大学医学院为第一作者单位。本课题受到国家重点研发(2018YFA0801000)、广东省基础与应用基础研究基金(2023A1515010396)和广州市科技计划(202201010156)项目资助。